Arachnoididae
Duncan, 1889, p.158
nomen transl. Arachnoidinae
Genre type : Arachnoides Leske, 1778
Description succincte de la famille : test plat avec zone ambitale aplatie, pťtales ouverts. 4 gonopores, prťsence de secteurs obliques ŗ l'extťrieur des zones ambulacraires alternant des rangťes de tubercules et de pores accessoires.
![]()
![]()

A revision of the Echinoidea from the Australian Tertiaries, p.158
 |
![]()
 |
|
I. Arachnoides placenta Ag. Tab. 21, fig. 35-42.
On reconnaÓt au premier abord cette espŤce ŗ sa forme lenticulaire, ŗ ses bords trŤs amincis, ŗ son profil subconique et ŗ son anus supra-marginal. J'ai long-temps partagť l'opinion des naturalistes qui l'envisagent omme congťnŤre des vrais Echinarachnius que nous venons de dťcrire dans le chapitre prťcťdent. Mais une ťtude plus approfondie des dťtails de son organisation m'a appris que les diffťrences qui les distinguent sont trop profondes pour que l'on puisse se refuser ŗ y voir deux types indťpendans. En effet, si l'A. placenta tient aux Echinarachnius par se forme circulaire, par ses pťtales ouverts et mÍme trŤs ťvasťs ŗ leur extrťmitť, et par la position de l'ouverture anale, qui est presque marginale, il s'en ťloigne par d'autres caractŤres peut-Ítre bien plus importans : et d'abord les sillons de la face infťrieure ne sont pas de mÍme nature, ils ne se ramifient point dans leur trajet du bord ŗ la circonfťrence, et, malgrť toute la peine que je me suis donnťe, je n'ai pu y dťcouvrir la moindre trace de pores microscopiques ; c'est une bande lisse et dťprimťe qui se continue ťgalement sur la face supťrieure jusqu'ŗ la rosette apiciale. En second lieu, la structure du test est trŤs diffťrente : au lieu d'Ítre massif, comme dans la plupart des Echinarachnius, il est au contraire trŤs mince et revÍtu d'une granulation toute particuliŤre, remarquable surtout par l'arrangement variable des tubercules. Sur les aires ambulacraires, les tubercules spinifŤres forment des sťries rťguliŤres obliques, sťparťes par de petites lignes en apparence continues (fig. 39), mais qui, examinťes sous un plus fort grossissement (fig. 39 a), se montrent composťes de tubercules miliaires conformťs de la mÍme maniŤre que les tubercules spinifŤres, c'est-ŗ-dire mamelonnťs et perforťs, quoique d'une petitesse extrÍme. Cette disposition des tubercules est la mÍme ŗ la face infťrieure, quoique moins apparente ŗ l'oeil nu. Les tubercules des aitres interambulacraires sont loin de prťsenter cette rťgularitť ; iles sont rťpartis sans ordre apparent ŗ la surface du test, et les tubercules sont entourťs de tubercules miliaires comme dans les genres prťcťdens. Cependant l'on remarque que ces derniers (les tubercules miliaires) sont proportionnellement moins nombreux (fig. 37 a). Un troisiŤme caractŤre qui distingue cette espŤce des Echinarachnius, c'est la prťsence de cinq pores gťnitaux, tandis qu'il n'y en a que quatre dans ces derniers. Ces pores occupent leur place habituelle au sommet des aires interambulacraires. Les pores ocellaires au nombre de cinq et ŗ peu prs aussi grands que les pores gťnitaux, sont placťs au xommet des pťtales ambulacraires et forment avec ces derniers un cercle rťgulier autour du corps madrťporiforme, qui est composť, comme ŗ l'ordinaire, d'une agglomťration de petits tubes capillaires invisibles ŗ l'oeil nu, mais trŤs distincts ŗ la loupe (fig. 41). Les ambulacres ont une forme trŤs particuliŤre ; ils ressemblent ŗ des triangles allongťs et ťtranglťs sur le milieu des flancs ; et comme ils s'ťvasent considťrablement, de maniŤre ŗ affecter ŗ peu prŤs les contours des aires interambulacraires dans d'autres genres, il est arrivť qu'on les a confondus avec ces derniŤres, et prťtendu que l'anus s'ouvrait dans un ambulacre. (Voyez la fig. 39 qui reprťsente un pťtale grossi). Les zones porifŤres ne se maintiennent comme zones continues que jusqu'ŗ la moitiť de la largeur du test ; passť cette limite, les pores n'existent plus que par paires isolťes que l'on poursuit jusqu'ŗ la circonfťrence. Les sillons transverses qui unissent les deux rangťes de pores d'une mÍme zone sont trŤs marquťs. Les pores eux-mÍmes sont ronds et trŤs uniformes. La structure de l'intťrieur justifie aussi sous plusieurs rapports la distinction que nous avons ťtablie entre ce genre et les Echinarachnius. A part les rťseaux ou piliers calcaires qui occupent les bords, on ne remarque ŗ la face interne du test d'autres aspťritťs que les auricules ; mais celles-ci sont d'une forme trŤs particuliŤre, car elles se terminent par deux petits bourrelets sťparťs par une ťchancrure trŤs profonde. L'appareil masticatoire est ausi grÍle que celui des Echinarachnius est massif : les m‚choires reprťsentent un compas trŤs ouvert (fig. 42 a, b) ; leur charpente est trŤs mince, hťrissťe de fines lames calcaires, sur les bords ; (voyez les fig. 42 c et d qui reprťsentent le profil d'une m‚choire vue par derriŤre et de cŰtť). Il existe en outre, ŗ la face supťrieure des m‚choires, une crÍte assez saillante qui s'ťlŤve de chaque cŰtť du sillon dentifŤre (fig. 42 b, c, d). La face infťrieure (fig. 42 a) n'est hťrissťe que de trŤs petites lames calcaires bien moins ťlevťes que celles de la face supťrieure. |
C'est ŗ ma connaissance la seule espŤce oý l'articulation des deux demi-m‚choires sur la ligne mťdiane soit distincte. Elles se dťsarticulent mÍme trŤs facilement, et il est alors facile de se convaincre que la demi-m‚choire droite est pourvue d'une petite carŤne longitudinale articulťe dans un sillon correspondant de la demi-m‚choire gauche. La bouche est centrale et circulaire, entourťe d'une rosette buccale peu distincte, mais qui paraÓt composťe de dix plaques, dont celles qui correspondent aux aires ambulacraires sont les plus ťtroites (fig. 36). J'ai reprťsentť (fig. 40) le pourtour de la bouche sous un grossissement de 1 1/2 diamŤtre, pour faire voir que les sillons qui viennent ici aboutir ŗ la bouche sont rťellement dťpourvus de pores microscopiques. Dans l'exemplaire qui a servi de modŤle ŗ la fig. 40, l'on voit quelques soies des bords des aires interambulacraires converger vers le centre de l'ouverture buccale, ŗ peu prŤs comme dans les Astťries. Une autre particularitť, c'est que dans l'enceinte de la rosette buccale, on ne remarque aucune diffťrence entre les tubercules des aires ambulacraires et ceux des aires interambulacraires ; ils ne reprennent leur disposition respective qu'en dehors de cette rosette. L'anus, comme nous l'avons dit plus haut, est supra-marginal, circulaire et paraÓt percť d'arriŤre en avant. Les soies sont trŤs fines, mais il ne paraÓt pas qu'elles diffŤrent beaucoup dans leur structure de celles des autres genres. Examinťes au microscope, elles se montrent hťrissťes de carŤnes fortement dentelťes ; celles de la face supťrieure sont clavellťes ; celles de la face infťrieure plus ou moins cylindriques. M. Forbes en a donnť une figure grossie dans son Histoire des Echinodermes anglais, pag. 181. Le Scutella Porpita des auteurs est bien certainement un jeune de l'Arachnoides Placenta ; je m'en suis assurť par la comparaison que j'ai faite des exemplaires du Musťe de Paris. Il m'est dŤs lors dťmontrť que l'espŤc fossile que M. Ch. DesMoulins a signalťe dans ses Tableaux synonymiques sous le nom de Cassidulus Porpita et que je ne connais pas, ne saurait avoir pour synonymes les citations relatives au Scutella Porpita des auteurs que M. DesMoulins lui rapporte ; car il m'est impossible de penser que M. DesMoulins ait pu prendre un jeune Arachnoides Placenta pour une espŤce du genre Cassidulus. C'est par erreur que l'on a cru cette espŤce originaire de l'Ocťan austral ; elle habite en rťalitť les mers du Nord. M. Forbes au moins assure que l'exemplaire qu'il a dťcrit et figurť dans son ouvrage a ťtgť recueilli par M. le professeur Jameson ŗ l'Óle Toulah, l'une des Óles Shetland ; mais c'est ŗ tort qu'il la confond avec l'espŤce du Canada de la collection de M. Lyell, qui est l'Echinarachnius parma, ainsi que nous l'avons dit plus haut en traitant de cette espŤce. Les originaux de mes figures m'ont ťtť communiquťs par MM. Valenciennes et J. E. Gray.
planche 21 (extrait)
|

Arachnoides placenta (Linnť,1758) - PlťistocŤne, Australie, 27 mm |
,%20apical,%20Pleistocene,%20Australie,%2027%20mm.JPG) |
|
,%20oral,%20Pleistocene,%20Australie,%2027%20mm.JPG) |
|
Genus Fellaster n. gen.
Fellaster may be differentiated from Arachnoides by the lack of
the periproctal groove and marginal notch; by the larger number of
interambulacral plates on the oral surface ; and by the smaller combed
areas. The type species was confused with Arachnoides placenta for
many years. To date, it has been authentically recorded only from New
Zealand, although it has The type species is strictly littoral, according to Mortensen. It has been recorded (Fell, 1953, p. 247) from the Pliocene of New Zealand. |
![]()
|
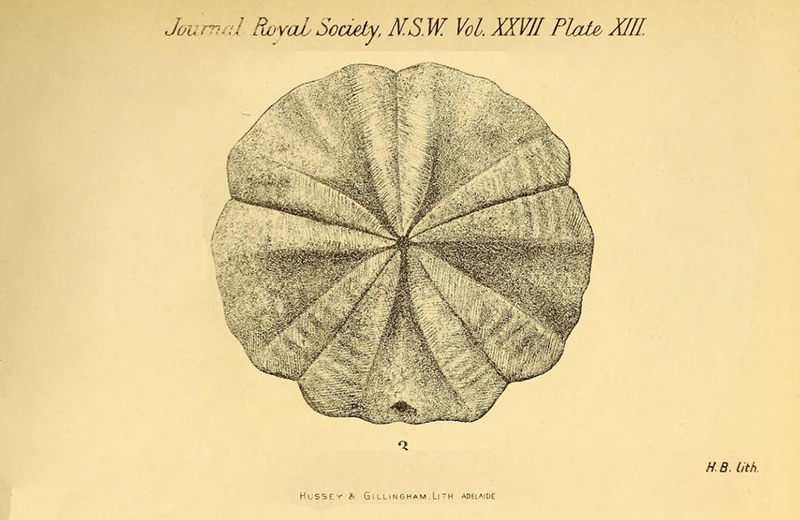
Arachnoides incisa, spec. nov., Pl. xiii., fig. 3.
Test very flat, rising slightly towards the apical disk, the longitudinal and transverse diameters are approximately equal; the apical disk is slightly in front of the centre. The ambitus is sharp and incised at the end of each interambulacral groove between which it is undulose. The ambulacre are slightly sunken and abruptly declivous at the sides, they occupy about an equal space with the interambulacra. The poriferoua zones reach about two-thirds way to the ambitus. The ornementation in the inter-ambulacre is obliquely banded and minutely granuler; the granulations are without order in the ambulacral areas. The periproct is supramarginal, with a concave depression between it and the ambitus, which is here slightly incurved. The actinal ares is flat. Antero-posterior diameter 54.4, transverse diameter 56.5, height 6.5 millimetres. Localities:óMiocene: Red Bluff and beyond Meinga, Gippsland Lakes, (three exemples). The incised ambitus, which mort markedly separates this fossil from the two known living species, recalls Monostychia from ail species of which it is separable by the superior position of the periproct. |
Except A. zealandiae, Gray, which dates bock to the Newer Tertiary of New Zeeland, this is the first occurrence of an extinct species of the genus ; A. conica, Hutton, of the Oamuru formation (=Eocene) in New Zealand, is a Monostychia. planche xiii (extrait)
|
 |
 |
|
Genre : fossulaster Lambert & Thiery Test dťprimť, ŗ pťtales indistincts et cloisons inconnues ; pťriprocte arrondi, supŤre et une large dťpression antťpťristomienne (marsupiale ?). Type : F. Halli Lambert, de l'EocŤne d'Australie, 1590 bis, p. 140, fig. se distingue du Eoscutum Morgani par son rostre postťrieur et la dťpression antťpťristomienne. |
![]()
confer supra
Fossulaster halli Lambert & Thierry, 1925 - OligocŤne, Australie du Sud, 10 mm spťcimen ♀ |
 |
 |
Fossulaster halli Lambert & Thierry, 1925 - OligocŤne, Australie du Sud, 9 mm spťcimen ♂ |
 |
 |
Fossulaster halli Lambert & Thierry, 1925 - MiocŤne, Gambier Limestone, Murray River, Australie du Sud, 11 et 9 mm spťcimen ♂ |
,%20apical,%20Miocene,%20Gambier%20limestone,%20Murray%20River,%20Australie%20du%20Sud,%2011%20mm.jpg) |
,%20oral,%20Miocene,%20Gambier%20limestone,%20Murray%20River,%20Australie%20du%20Sud,%2011%20mm.jpg) |
,%20apical,%20Miocene,%20Gambier%20limestone,%20Murray%20River,%20Australie%20du%20Sud,%209%20mm.JPG) |
,%20oral,%20Miocene,%20Gambier%20limestone,%20Murray%20River,%20Australie%20du%20Sud,%209%20mm.JPG) |
Fossulaster halli Lambert & Thierry, 1925 - Miocene superieur, Port Campbell, Nuna beach, Portland, Victoria, Australie, 4 mm spťcimen ♂ |
 |
 |
|
Monostychia Laube. Der KŲrper ist flach schilfŲrmig, lšnger als breit. Der Rand ist ziemlich scharf vorn abgerundet, hinten in ein kurzes breites Rostrum ausgezogen, gegenŁber jedem Petaloide une in der Mitte des Rostrums einmal eingekerbt. Diese Kerbungen sind nicht alle gleich. Die Kerben des unpaaren Petaloides und des vorderen Paares sind gleich une schwšcher als die tiefen Kerben des hinteren Petaloidenpaares, welche stumpfe Winkel bilden, deren einen Schenkel das Rostrum bildet. Der Kerbe in Mitte das Rostrum bildet. Der Kerbe in Mitte des Rostrums ist grŲsser als die vorderen, kleiner als die hinteren Randkerben. Die Oberseite der Schale ist schwach gawŲlbt, nach hinten flacher und fast in gerader Linie, nach vorn steiler und in etwa gekrŁmmter Linie abfallend. Der Scheitel liegt etwas vor der Mitte und zeigt vier Genitalporen, deren vorderes Paar nšher zusammensteht als das hintere. Die Petaloidien reichen etwas bis Łber die Mitte der Scheibe und sind vorn weit geŲffnet. Die Ambulacralzonen zeigen schršgstehende durch scharfe Rinnen conjugirte Porenpaare. Die šussere Pore ist geschlitzt, die innere rundlich, man zšhlt je nach dem Alter 20-30 Paare. Die šussere Porenlinie ist stšrker als die innere gekrŁmmt, oben ist die Porenzone fast geschlossen. Die Interporiferenzone ist fast doppelt so breit als die eine Poriferenzone, sie ist glatt une ragt Łber die etwas vertieften letzteren merklich hervor, sie ist spitz dreieckig, mit kaum gebogenen Ršndern mit der Spitze gegen den Scheitel gelegen. Die Interambulacralfelder sind zwischen den Petaloidien ganz schmal, nehmen unter diesen rasch an Breite zu, und sind am Rande selbst etwas breiter als die Ambulacralfelder. Die Oberseite ist mit feinen regellos stehenden Wšrzchen bedeckt. Die Unterseite ist ganz flach, bei einselnen Individuen gegen dit Mitte schwach ausgehŲhlt. Das Peristom ist central, rund, bei jungen Exemplaren fŁnfseitig. Vom Peristom gegen den Rand verlaufen fŁnf einfache, tiefe Ambulacralfurchen, die unpaare gerade, die paarigen schwach gegen einander gekrŁmmt, welche am Rande genau in die Kerben einmŁnden, und Łber diese hinaus auf der Oberseite Łber das Ambulacralfeld bis auf die Interporiferenzone fortsetzen und erst in der Nšhe des Scheitels ganz verschwinden. Das Periproct liegt am hinteren Rande dicht unter der Mittelkerbe im Rostrum, eine schwache doch merkliche ErhŲhung zieht sich vom Peristom zum Periproct hin. Die Unterseite ist mit grŲheren ebenfalls regellos stehenden Warzen bedeckt. |
Ob Kiefern vorhanden sind liess sich nicht beobachten, wohl aber zeigt das innere unter den Interambulacralfeldern je zwei bie ovn die Mitte reichende Lšngstršger, von welchen zahlreiche QuerstŁtzen unter die Ambulacralfelder abgehen. Vorstehend beschriebenes Geschlecht hat von oben besehen sehr viel ńhnlichkeit mit Scutella, allein man braucht den KŲrper nur umzukehren, um sofort zu erkennen, dass die Abwesenheit anostomosirender Ambulacralfurchen diesem widerstreiten. Die einfachen Ambulacralfurchen gestatten nicht einmal die Einreihung des Seeigels in die Zunft der Scutelliden, sondern weisen ihn in jene der Laganen. Unter diesen kŲmmt als nšchstverwandtes Geschlecht die in dem australischen Ocean lebende Arachnoides Klein in Betracht. Ausser der Form der weit getrennten Poriferenzonen, haben sie beide nochden Kerbrand und die einfachen, vom Mund bis fast zum Scheitel reichenden Ambulacralfurchen gemein. Allein ausserdem, dass bei Arachnoides die Form kreisrund, hier lšnger als breit ist, dass bei ersterer die Poren regelmšssig in Zeilen, hier regellos stehen, liegt bei ersterer das Periproct Łber dem Rande, bei letzterer unter demselben, worin ein wesentlicher Unterschied besteht, eben so wie darin, dass jene fŁnf, diese nur vier Genitalporen hat. Vom Genus Rumphia Desor unterscheidet sich Monostychia wesentlich durch das Vorhandensein eines inneren TraggerŁstes. Von diesem Geschlechte ist bis jetzt eine einzige Art bekannt geworden, welche demnach den Typus ausmacht, es ist dies...
|
![]()
,%20apical,%20Miocene%20inferieur,%20Victoria,%20Australie,%2020mm.JPG) |
|
,%20periprocte,%20Miocene%20inferieur,%20Victoria,%20Australie,%2020mm.JPG) |
,%20oral,%20Miocene%20inferieur,%20Victoria,%20Australie,%2020mm.JPG) |
![]()
Monostychia sp. "Knight shield" type |
NOTA : Cette forme particuliŤre et rťcurrente chez Monostychia n'est pas encore documentťe. Elle est apparue au milieu des annťes 90, au sein d'une donation au Museum de Victoria (coll. Foster) et lors de l'examen/cataloguage de cette derniŤre. Un travail de rťvision serait en cours, incluant cette forme, mais fait de maniŤre non continue. Je tente de rentrer en contact avec la personne en charge de ce travail afin de savoir si une publication officielle est prťvue ou non, et ŗ quelle ťchťance. Pour l'heure, ce morphotype (?), espŤce (?) est traitť en nomenclature ouverte sous le nom "type Knight Shield" en attendant la publication valide.
Monostychia sp. "Knight shield" type - OligocŤne supťrieur (Janjukian), "Wool Bay", Pťninsule de York, Australie du Sud, 20 mm |
 |
 |
Monostychia sp. "Knight shield" type - OligocŤne supťrieur (Janjukian), "Wool Bay", Pťninsule de York, Australie du Sud, 23 mm |
 |
 |
|
Phylogenetic analysis of the fossulasterid echinoids (Order Clypeasteroida) from Australia, p.229 EspŤce type Scutellina morgani Cotteau, 1891 (dťsignation originale) Extension stratigraphique (bibliographique, non vťrifiťe) : OligocŤne supťrieur Syn. |
![]()
Philipaster occultoforma (Irwin,1995) - MiocŤne, Batesford quarry, Batesford, victoria, australie, 10 mm |
,%20apical,%20Batesford%20quarry,%20Batesford,%20victoria,%20australie,%20MiocŤne,%2010%20mm.jpg) |
,%20amb,%20Batesford%20quarry,%20Batesford,%20victoria,%20australie,%20MiocŤne,%2010%20mm.JPG) |
,%20oral,%20Batesford%20quarry,%20Batesford,%20victoria,%20australie,%20MiocŤne,%2010%20mm.jpg) |
|
Genre Prowillungaster Wang,1994 Phylogenetic analysis of the fossulasterid echinoids (Order Clypeasteroida) from Australia, p.229 EspŤce type Prowillungaster major Wang, 1994 (dťsignation originale) Extension stratigraphique (bibliographique, non vťrifiťe) : OligocŤne supťrieur ŗ MiocŤne infťrieur |
![]()
Prowillungaster major Wang,1994 - MiocŤne, Murray River, Australie du Sud, 9 mm |
,%20apical,%20Mannum%20formation,%20Miocene,%20Murray%20River,%20Australie%20du%20Sud,%209%20mm.JPG) |
,%20oral,%20Mannum%20formation,%20Miocene,%20Murray%20River,%20Australie%20du%20Sud,%209%20mm.JPG) |
|
Genus Scutellinoides n. gen. Type species: Scutellina patella Tate (1891) = Scutellina morgani Cotteau (1891), not S. patella Tate of Hall (1908). Externally similar to Scutellina except for supramarginal periproct; apical surface depressed conical; oral surface slightly concave ; margin moderately thin, rounded ; petals moderately well developed, extending nearly three-fourths distance to margin, somewhat open; pores rounded, not conjugated; apical system and peristome central; periproct supramarginal, distant about two pairs of plates from ambitus; basicoronal interambulacral plates apparently small; interambula≠cral areas not connecting on oral surface. Miocene, Australia. Examination of specimens from the Tate Collection shows that this species lias separate auricles, marginal concentric supports, and an inner group of radial interambulacral supports, characteristics which clearly indicate that it is a member of the arachnoidid subfamily Ammotrophinae rather than the Fibulariidae. Its supramarginal periproct is distinctive within this group. In the available specimens it cannot be determined for certain whether there are any post-basi≠coronal interambulacral plates on the oral surface, but the interambulacra do not appear to pass onto the oral surface. On the oral surface |
they arc
separatcd by at least one and perhaps two or more pairs of enlarged
ambulacral plates. The peripheral concentric supports are not as
extensively developed as in Ammotrophus, but the inner radial
supports are. The combed areas are certainly not wcll de≠veloped, but
on the oral surface there arc a few oblique rows of secondary
tube≠feet pores that may represent the combs. The primary tubercles
are well developed, prominent, and perforate. Examination of specimens in the British Museum (Natural History) of the species with a marsupium described by Hall (1908), and referred to Scutellina patella Tate, shows that they represent a distinct species which Lambert and Thiťry (1925, p. 577) named Fossulaster halli and for which they proposed the genus Fossulaster. Hall's species differs from Tate's in that its shape is more elongate, it has internally only two pairs of inner radial supports rather than five pairs, and the female has a marsupium. |
![]()
|
Scutellina
patella, spec. nov.
Test large for the genus ; subcircular to subdecagonal in outline ;
regularly conic. |
Tubercles uniform, small, and perforate, rather wide apart, surrounded
by scrobiculate ornament. Dimensions of two examples :óLongitudinal diameter, 24, 22 ; transverse diameter, 22.5, 21; height, 7.5, 6. Localities.óCalciferous sandstones of the River Murray Cliffs (connnon); white polyzoal limestone at Mount Gambler; polyzoal sands, ]Muddy Creek. By its supra-marginal periproct this new Scutellina is related to S. supera, Agassiz; but its circular outline and high summit distinguish it ; it is, moreover, very much larger. |
Scutellinoides patella (Tate,1891) - MiocŤne, Murray River beds, Sm quarry, North Youghusband Store, Australie du Sud, 10 mm |
,%20apical,%20Miocene,%20Murray%20River%20Beds,%20Sm%20quarry,%20North%20of%20Youghusband%20Store,%20Youghusband,%20Australie%20du%20Sud,%2010%20mm.jpg) |
,%20oral,%20Miocene,%20Murray%20River%20Beds,%20Sm%20quarry,%20North%20of%20Youghusband%20Store,%20Youghusband,%20Australie%20du%20Sud,%2010%20mm.jpg) |
|
Genus willungaster gen. nov. Type species. Willungaster scutellaris sp. nov. Diagnosis. Moderate sized fossulasterids, with rounded outline. Interambulacra extremely reduced to narrow columns extending only about two-thirds of the distance to the ambitus ; periproct supramarginal and in large specimens located at the termination of the posterior interambulacral column. Ambulacral petals well defined, open, with numerous slightly conjugate pores. Ambulacral food grooves not developed. Apical system with four genital pores. Girdle of separate auricles ; inner supports paired and interradial ; concentric supports developed at margin only. Females with a marsupium developed on the adoral surface to the anterior of the peristome. |
Remarks. Willungaster differs from other fossulasterids in its strongly reduced interambulacral columns. In the other genera (Fossulaster, Scutellinoides, 'Scutellina' patella Tate) the interambulacra extend to the ambitus and the supramarginal periproct is contained within the posterior interambulacral column. Indeed, in Willungaster the interambulacra are more reduced than in any other echinoid genus. |
![]()
|
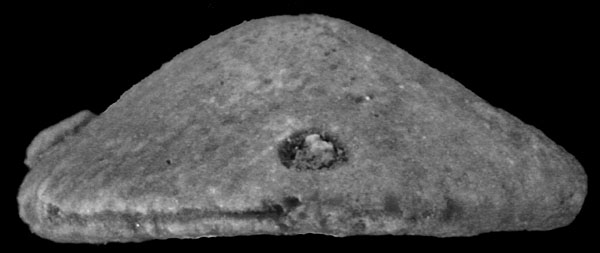
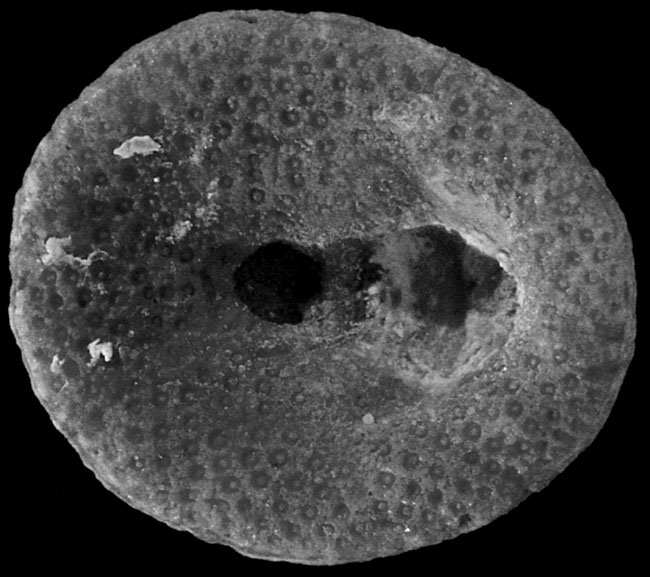
Willungaster scutellaris sp. nov. Material. Holotype P27951 (♀), paratypes UNE 11675 (♂), 11766 (♀), 1767, 12006 (♂), 12007 (♂), 12008 (♂), and 141 other specimens (62♂, 71♀, 8 indet.) from Edithburg, Yorke Peninsula, S.A. from a Janjukian or Longfordian (Upper Oligocene of Lower Miocene) horizon in the Port Vincent Limestone. Description. Test rounded in outline, in lateral view gently arched to reach maximum height at the apical system. Periproct rounded, just supramarginal, and maintaining this position during growth ; in small specimens (Pl.131, fig.5) located between two enlarged posterior ambulacra ; in mature specimens abutting against the same plates and the termination of the posterior interambulacral column (Pl. 131, fig. 7 ; text-fig. 8a). Interambulacral columns narrow, in small specimens reaching about half-way to the ambitus ; in larger ones extending up to three-quarters of this distance (Pl.131, fig.7). Apical system somewhat posterior, small and with four genital pores. Ambulacra with well-developed open petals with slightly conjugate rounded pores mounted in the horizontal sutures. |
Adoral surface flattened in males, in females with a marked depression to the anterior of the rounded peristome. Plating of adoral surface consisting of three circlets of enlarged ambulacral plates, with a supplementary plate in column IIIa, present in both males and females (text-fig. 8b, c). Marsupium of females floored by the supplementary plate and plates of column III second from the peristome. Specimens with a marsupium usually more arched in lateral profile (cf. Pl. 131, figs. 1 and 2). Basicoronal interambulacral plates not visible. Surface of test covered with small, closely spaced, sunken tubercles, except within the marsupium where the test is smooth or sparsely granulated. Interior of test with small separate auricles, and five well-developed paired interradial pillars (Pl. 133, fig.9). Up to two series of concentric supports within the ambital margin of the test.
|
Willungaster scutellaris Philip & Foster, 1971 - OligocŤne, Pťninsule de Yorke, Australie, 14 mm spťcimen ♀ montrant la dťpression incubatrice |
 |
 |
Willungaster scutellaris Philip & Foster, 1971 - OligocŤne supťrieur, Port Willunga formation, Pťninsule de Yorke, Australie du Sud, 11 mm spťcimen ♀ montrant la dťpression incubatrice |
 |
 |
 |
 |
Willungaster scutellaris Philip & Foster, 1971 - OligocŤne, Pťninsule de Yorke, Australie, 11 mm spťcimen ♂ |
 |
 |
Willungaster scutellaris Philip & Foster, 1971 - MiocŤne, Mont Gambier Lmst, Mt Gambier, Australie du Sud, 9 mm spťcimen ♂ |
,%20apical,%20Gambier%20limestone,%20Miocene,%20OB%20Flat,%20Mont%20Gambier,%20Australie%20du%20Sud,%209%20mm.JPG) |
,%20oral,%20Gambier%20limestone,%20Miocene,%20OB%20Flat,%20Mont%20Gambier,%20Australie%20du%20Sud,%209%20mm.jpg) |