
Systématique
![]()
![]()
I. - L'espèce, c'est quoi ?
On peut grosso-modo percevoir la notion d'espèce au travers de trois reflets de la réalité. Nous utilisons sciemment le terme de reflet car la notion d'espèce est une pure création de l'esprit humain, résultant de son ardent désir depuis l'antiquité pour tout hiérarchiser, classer, nommer, bref : tout dominer. En même temps, le terme de reflet est un peu exagéré puisqu'une des trois conceptions de l'espèce décrites ci-dessous est purement naturelle. En tous cas, si e n'est les abus de langage qui sont régulièrement commis, on relègue alors cette notion à une entité purement intellectuelle, un objet de l'esprit tel qu'ont pu l'utiliser les philosophes depuis le siècle des lumières. Et quoi qu'il en soit, il ne fait aucun doute que le temps aidant, la notion d'espèce évoluera et que l'on trouvera encore bien des choses pour la mieux caractériser encore. Nous aurons l'occasion de revenir en détail dans un article ultérieur sur le concept d'espèce, qu'il soit biologique, opérationnel ou évolutif. La place et le temps manquent ici, ne serait-ce que pour aborder ces concepts.
Pour en revenir au sujet qui nous préoccupe, nous allons considérer rapidement trois notions relatives à l'espèce. Des notions plus que de réels concepts acceptés (voir paragraphe précédent), car il nous faut bien à un moment parler de taxa, et que l'espèce à un moment ou à un autre va ressurgir dans le discours sur la systématique.
L'espèce biologique : Il s'agit d'un groupe de spécimens qui partagent le même patrimoine génétique. Le caractère état naguère exprimé par la notion d'interfécondité. L'utilisation récente de la génétique venant appuyer la notion d'interfécondité. L'espèce biologique, on s'en rend compte tous les jours, représente à nos yeux un groupe d'individus qui entrent dans un champ de variations individuelles, mais dont certains caractères sont communs. Statistiquement, la représentation de ces caractères communs s'inscrit dans une courbe normale, dite courbe de Gauss, ou "en cloche". Les individus les plus nombreux sur cette courbe seront donc ceux (pour un caractère donné), se rapprochant le plus du point médian (le caractère moyen). Pour représenter cette espèce, on définit un holotype dont les caractères propres à son espèce se rapprochent le plus possible des caractères moyens. De part et d'autre de ce caractère moyen sur la courbe, des individus se répartissent tout le long de la courbe de Gauss. Plus on s'éloigne dans un sens ou dans un autre, moins le nombre de spécimens est important : l'espèce est donc bien caractérisée. Il arrive cependant qu'à l'intérieur d'une espèce biologique on observe des morphologies très différentes. Elles peuvent s'expliquer par des facteurs sexuels (dimorphisme sexuel), géographiques, environnementaux. Il y a donc polymorphisme, avec pour conséquence, l'échantillonnage possible de plusieurs populations distinctes à l'intérieur de la même espèce : on parle de polymorphisme et par conséquent de polytypisme (plusieurs types).
L'espèce dite typologique : C'est la notion la plus ancienne. Elle est caractérisée par un holotype fondé UNIQUEMENT sur des considérations morphologiques. Le champ de variation de l'espèce typologique est donc hautement variable en fonction de l'observateur ou du descripteur (notion de subjectivité). Une bonne illustration de cette subjectivité est l'explosion du nombre de création d'espèces chez les Céphalopodes fossiles, par les auteurs du début du XXème siècle.
L'espèce en paléontologie : Elle est proche de l'espèce typologique, sauf lorsqu'elle est définie sur une population d'individus testée comme normale. Lorsqu'une espèce est fondée sur quelques spécimens (voire un seul !) on parle d'une espèce purement typologique. Les espèces fondées sur des populations s'écartent un peu du caractère purement typologique, puisque théoriquement si cette population est correctement échantillonnée, elle contient : des jeunes, des adultes, des séniles, des morphes extrèmes. Des outils statistiques permettent de tester les populations recueillies pour voir si elles s'ajustent sur une courbe de Gauss et par conséquent pour voir si la récolte, l'échantillon sont bien le reflet d'une population telle qu'on pourrait la trouver spontanément dans la nature. On voir donc clairement l'incidence par exemple si on ne recueille que les plus beaux spécimens, si on néglige les petites formes, etc... Quoi qu'il en soit tout est basé ici sur la morphologie et on peut avoir plusieurs espèces typologiques alors que la réalité de l'espèce biologique n'en a conçu qu'une seule.
Afin d'écarter le plus possible ce genre d'artefact, il est utile de considérer les échantillonnages multiples de manière à recouvrir le plus largement possible les champs de variation possibles d'une espèce : intra-populationnel (dimorphisme sexuel par ex.), temporel (passage progressif depuis le bas d'un gisement, vers une autre forme dans le haut du gisement), spatial ou géographique (au travers de plusieurs gisements éventuellement représentatifs de paléo-environnements différents) Pour faire simple on a donc plus intérêt à désigner une (des) population(s) comme types : désigner plusieurs holotypes moyens. Dans ce domaine aussi des outils existent pour opérer des regroupements ou des discriminations. Les statistiques avec les analyses multivatiées, ou bien la morphométrie géométrique (pour ne citer que ces deux là, supports sur lesquels des articles suivront).
II. - L'espèce et la systématique
Nous voilà bien... avec nos... espèces ! Qu'en fait-on ? Mettons nous juste une étiquette avec nos spécimens maintenant baptisés, ou tentons nous d'aller un peu plus loin ? Allez, c'est parti !
La systématique vise à décrire les organismes en utilisant les relations inter-spécifiques et leur degré de proximité. Outre son objet, elle regroupe les disciplines de la taxinomie, de la nomenclature, la classification pure. La taxinomie classe (hiérarchise) les regroupements opérés. La nomenclature est utilisée par la taxinomie pour nommer les hiérarchies définies par elle même. La classification, quant à elle, s'occupe d'insérer un groupe dans le système ainsi hiérarchisé. Rappelons ici l'existence de l'ICZN (International Code of Zoological Nomenclature) qui présente toutes les règles de la nomenclature in extenso, cet organisme les gère, les publie, les actualise. Sachez cependant qu'il est souverain, mais n'impose pas (ce n'est pas une loi, mais un code). Ainsi on se retrouve parfois avec des bizarreries nomenclaturales dans les noms d'espèce car certains n'ont pas suivi (ou pas voulu suivre) les recommandations de ce code. C'est bien dommage, car pour être bien compris, il est nécessaire que tous parlent dans la même "langue".
On regroupe tout d'abord certaines espèces au sein de genres, car ces espèces entretiennent de fortes relations communes. Le genre est donc le niveau hiérarchique immédiatement supérieur à l'espèce (si on exclut les catégories infra-génériques intermédiaires : le sous-genre en l'occurence).
Beaucoup d'entre vous s'en sont rendu compte : la systématique évolue en permanence (d'où l'utilité d'user d'une langue nomenclaturale correcte et de se reporter fréquemment au code de l'ICZN lorsque l'on "commet" un acte nomenclatural)..
Au travers du vocable de systématique, on distingue plusieurs disciplines (ou plutôt des "Ecoles") : la phénétique, la phylogénétique, la systématique évolutive.
C'est ici que cela se corse... Nous allons détailler les grandes lignes de trois méthodes, tout en restant superficiels et sachant que nous n'en détaillerons sommairement qu'une seule.
III. - La phénétique
Elle est basée sur l'utilisation de l'ensemble des caractères discriminants présentés par le génotype, soit : le phénotype.
Cette systématique utilise les méthodes statistiques numériques afin d'examiner les caractères communs à plusieurs espèces. Son gros handicap est de donner le même poids aux différents caractères pris en compte, ce qui dans la réalité peut conduire à des résultats... bizarres, pour ne pas dire totalement aberrants. Les formes de sortie après traitement par cette méthode sont des arbres appelés phénogrammes (ou dendrogrammes). Afin d'illustrer le discours ci-dessus, voici un exemple comparant le singe, le cheval et le lézard à collerette. La méthode consiste à compter le nombre de caractères communs entre chaque espèce. Si nous considérons les caractères suivants :
-
caractère A = 5 doigts différentiés à la patte antérieure ;
-
caractère B = présence ou non d'une queue ;
-
caractère C = deux yeux
| espèce | caractère A | caractère B | caractère C |
| singe | X | X | X |
| lézard à c. | X | X | X |
| cheval | O | X | X |
D'après le tableau, le singe et le lézard à collerette ont chacun 3 caractères en commun. Par contre ce même singe n'a que deux caractères en commun avec le cheval (B et C). On en conclut que le singe est plus proche du lézard à collerette que du cheval. Sympathique, non ? Bien sûr l'exemple est fait par l'absurde, le nombre de caractères vicieusement peu élevé... Mais le problème vient surtout du fait que dans la réalité, les différents caractères n'ont pas la même importance. Or la phénétique ne tient pas compte de ce facteur. Le fait de considérer tous les caractères avec le même poids vient d'un louable souci : celui d'essayer de ne pas introduire de subjectivité. Cependant, une des qualités de la méthode finit par devenir un de ses plus gros défauts. Ce défaut peut être atténué, mais pas supprimé, en considérant un très grand nombre de caractères.
IV. - La systématique phylogénétique (cladistique)
On utilise l'évolution biologique par descendance en considérant les modifications. Le but est de trouver les degrés de parenté entre les différents groupes, de trouver leur ascendant commun. On met en relation des groupes grâce à des caractères que l'on qualifie d'un point de vue évolutif. L'intérêt de la méthode est de qualifier LA LOGIQUE entre les caractères utilisés : on analyse donc non seulement les caractères, mais aussi leur distribution. Le type de troncs construits grâce à cette systématique est nommé : cladogramme.
On considère deux types de caractères (qualification des caractères) : PLESIOMORPHES (primitifs) du grec plesios = voisin et morphos = forme et APOMORPHES (caractères dérivés) du grec apo = éloigné et morphos = forme. Les filiations obtenues avec ces caractères sont caractérisées par trois grands cas :
-
Synapomorphie : partage d'un état apomorphe, (on parle alors d'un groupe monophylétique) plusieurs caractères apomorphes sont partagés par le groupe étudié ;
-
Symplésiomorphie (on parle alors d'un groupe paraphylétique, c'est-à-dire constitué de sous-groupes monophylétiques) : plusieurs caractères plésiomorphes sont partagés par le groupe étudié : ils ont en commu des caractères primitifs, ancestraux ;
-
Homoplasie (on parle alors d'un groupe polyphylétique) : les individus du groupe étudié se ressemblent de par le fait d'une convergence de forme adaptative. L'étude de groupes polyphylétiques permet parfois d'affiner la filiation, par de nouvelles observations ou à la lumière de caractères restés non étudiés.
C'est bien beau tout ça, mais on fait comment ?
Quels caractères ?
On choisit un groupe de référence extérieur (OUT ou OUTGROUP) qui théoriquement NE PEUT englober le groupe que l'on est en train d'étudier.
On choisit des individus dans le groupe que l'on va étudier (IN) pour déterminer leurs caractères homologues. Pourquoi faire ici un choix d'individus pour le groupe ? Parce que les outils utilisés (informatique) sont dépassés par les temps de traitement (temps machine) qui croissent de manière absolument affolante au fur et à mesure que l'on augmente le nombre d'individus choisis pour l'étude.
Subjectivité : les caractères choisis doivent impérativement traduire une homologie réelle entre deux caractères. On devrait donc plutôt parler d'un choix de caractères TRADUISANT UNE HOMOLOGIE, plutôt qu'un choix de "caractères" (toutefois, ce vocable est plus confortable pour le discours).
Dans quel sens sont orientés ces caractères ? (polarisation)
Les caractères homologues sont décidés comme appartenant à l'une ou l'autre des catégories :
-
Primitif (0) : hérité SANS modification aucune à l'intérieur d'un clade (et seulement dans ce clade) ;
-
Dérivé (1) : hérité AVEC transformation une seule fois entre deux entités à l'intérieur d'un clade (et seulement dans ce clade).
On part donc à la base sur différentes hypothèses d'évolution variant en fonction des caractères, car ces derniers dépendent de deux critères (dont l'un a été défini plus haut) :
Le critère OUT ou de OUTGROUP : l'Aïeul de cet outgroup (son ancêtre commun) est antérieur à l'ancêtre commun le plus ancien du groupe que l'on est en train d'étudier. Si une espèce appartenant à l'outgroup possède le caractère dont on recherche la polarisation et ce, sous une forme identique, ce caractère est dit VOISIN (plésiomorphie / primitif). Si il est au contraire éloigné (apomorphie), il sera qualifié de DERIVE (synapomorphie entre les deux caractères, voir plus haut). Remarques importantes : Par ancêtre ou aïeul, nous entendons ici une entité conceptuelle regroupant certains caractères, et non pas un individu, ou un groupe au sens physique du terme. Ainsi, par ancêtre, il faut comprendre qu'au sein de la similarité de caractères de la lignée, on parle bien du plus éloigné similairement parlant. Deux rameaux parallèles auront donc leur point de convergence en un seul point qui concentrera la plus proche proximité entre tous les caractères communs aux deux rameaux.
En ce qui concerne les qualificatifs des caractères (primitifs ou dérivés) : il ne faut pas leur attribuer un sens temporel, mais plutôt d'éloignement et de proximité (voir l'étymologie des termes synapomorphie et plésiomorphie plus haut dans le texte). Comme on l'a fait remarquer au début de cette partie sur le cladisme, c'est la logique qui régit cette forme de systématique. Donc les relations phylogénétiques exprimées par la méthode seront celles qui satisferont au mieux les hypothèses retenues, en terme de pure logique (principe de PARCIMONIE). Encore une fois, se basant sur la logique et en dehors d'un cadre temporel, on peut considérer les résultats comme raisonnables fonction des contraintes de départ. On pourrait dire : plausibles.
Le critère de l'ONTOGENIE : Le regard sur le développement d'un individu montre que : plus un caractère apparaît tardivement dans l'ontogénie, plus il est spécialisé et dérivé (apomorphie) ; plus il apparaît tôt dans le développement, plus il est primitif (plésiomorphie).
Quelques définitions de termes utiles pour nommer les différents groupes et leurs connexions :
-
Le clade : c'est un ensemble de spécimens regroupés à l'aune de leurs caractères homologues. Le clade au vu de ce qui a été dit précédemment est donc un groupe monophylétique bien défini : c'est le cas de l'OUTGROUP.
-
Homologie de filiation : définie par une parenté exclusive (relation logique de filiation). On dit donc de deux caractères qu'ils sont homologues s'ils sont présents chez deux spécimens ayant un ancêtre commun exclusif. On ne peut poser une homologie avec un seul caractère, puisque comme nous l'avons précisé au début de partie sur cette systématique : on qualifie la logique entre deux caractères.
-
caractères homologues : caractères reconnus chez deux individus ayant un ancêtre commun exclusif, donc pas de possibilité d'homologie si on parle d'un seul caractère (rappel)
Optimisation de la logique de l'arbre (cladogramme) : Les caractères homologues permettent de construire UN SEUL ARBRE, nommé ARBRE LE PLUS PARCIMONIEUX, car l'arbre le plus court est considéré comme le moins faux. Cela signifie que l'ensemble des individus ne sont qu'une partie agissante et dépendante par rapport au phénomène de la vie passée et présente.
La représentation graphique, précisions : On l'a vu, le cladogramme est censé représenter des liens de ressemblance logique entre individus vivants ou fossiles (pour ce qui nous concerne, puisque la démarche cladistique est capable de classer, comparer des objets inanimés : tout est ici affaire de logique). Sur la représentation graphique, les éléments support de comparaison sont généralement encadrés (ce sont les caractères).
Pour conclure provisoirement sur la cladistique, on peut dire que cette systématique compare les taxa. On définit un groupe comme étant un ensemble de caractères, chaque caractère présentant un état (polarisation). Les taxa sont considérés deux à deux (groupes frères), à l'aune des caractères présentant un état dérivé. Sa représentation graphique, le cladogramme n'est PAS un arbre évolutif, il constate et est donc critiquable et reproductible. Il ne permet par conséquent pas d'imaginer des scenari. Cette systématique rejette absolument les groupes extra-monophylétiques, seuls les clades reçoivent donc un nom.
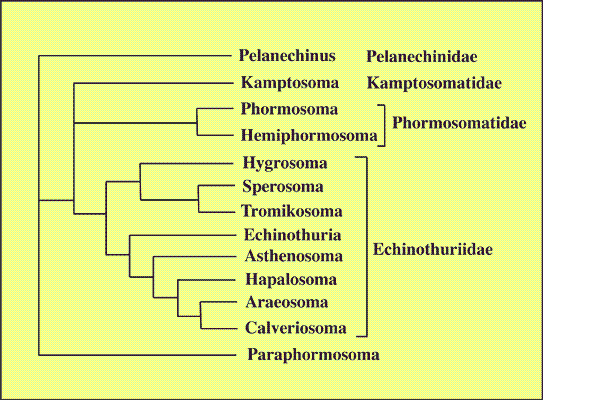
exemple n°1 de cladogramme pour l'ordre des Echinothurioida CLAUS,1880 (d'après SMITH A.B. (Ed.) 2003, The echinoid directory. World Wide Web electronic publicationK. http://www.nhm.ac.uk/palaeontology/echinoids [accessed 06 January 2005]).
|
|
|
Les caractères pris en compte : Apical disc Ambulacra Peristome Interambulacra Tuberculation Spines and pedicellariae Teeth
|
|
La matrice obtenue à partir de ces caractères (remarquer l'incertitude parfois dans la polarisation des caractères : "?") Pelanechinus 10000 01101 00100 00110 00000 00011 00? Kamptosoma 0?101 12000 00010 01000 01010 12000 010 Phormosoma 01001 02000 00000 00010 00111 00111 000 Hemiphormosoma 01001 02200 01000 00010 00111 00100 00? Paraphormosoma 00000 02110 00000 00010 00000 00001 00? Hygrosoma 00010 02000 00000 00010 00111 01001 101 Tromikosoma 01011 11100 01001 00000 00001 01010 10? Sperosoma 01010 11100 11001 00010 00001 01010 101 Echinothuria 10000 02200 01000 10010 00010 11??1 ??? Asthenosoma 11000 02200 01100 10010 10011 01001 001 Hapalosoma 10000 02100 01100 10011 10010 11010 011 Araeosoma 10010 02200 01100 10111 10001 11001 011 Calveriosoma 10010 02200 01100 10111 10011 11001 001
|
exemple n°2 de cladogramme pour les genres d'oursins Camarodontes (modifié d'après SMITH A.B.(1998),
| Dans ce type de cladogramme, la matrice des caractères et leur polarisation sont incluses dans le graphique lui même : la lecture est plus touffue, mais plus directe. Comparer avec l'exemple 1 ci-dessus. |
|
|
|
Les caractères pris en compte : (je vous engage à consulter cette note qui est reprise en bibliographie de cet article, afin de lire la discussion sur la polarité de chaque paramètre, illustrant parfaitement les propos ci-dessus) Symboles ouverts = caractères primitifs Symboles pleins = caractères dérivés Doubles barres = caractères présentant des états multiples pour lesquels un état (polarisation) est plus inclusif qu'alternatif. "?" = caractère avec polarisation inconnue caractères dérivés n°1-20 = liste ci-dessous 1. blade of globiderous pedicellariae compact and coalesced 2. primary tooth plate with distance from umbo to lateral edge (compare distance fram umbo to oral edge in the range 0,8-1,2 [circles] or 1,3+ [squares]) 3. primary tubercles weakly crenulate (half-filled circles) or noncrenulate (solid circles) 4. test with broad naked interradial and perradial bands adapically 5. globiferous pedicellariae with single poison glands 6. CLNP tooth system with oral lamellae that are branched or forked 7. globiferous pedicellariae with a single asymmetrical side tooth (circles) or no side teeth (squares) 8. CLNP tooth system with a comb of tines 9. stalk of globiferous pedicellariae solid (squares) or tubular (circles) 10. apical disc with oculars I and V insert 11. larvae with a basket-type skeleton 12. globiferous pedicellariae with stalk glands 13. buccal notches sharp and deep, extending level to the fourth ambulacral plate 14. ambulacra with a primary tubercle only on every second or third compound plate 15. ambulacral pores arranged in two (squares) or three (circles) discrete columns on each compound plate 16. test elliptical in outline 17. ambulacra polyporous 18. polyporous ambulacra with eight or more component elements 19. globiferous pedicellariae with muscular neck 20. sutural pits present on coronal plates |

IV. - La systématique évolutive
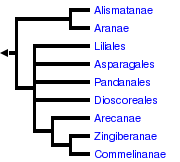
Elle construit des représentations nommées : phylogrammes. Ces derniers représentes les liens, les parentés que l'on a pu dédure en considérant les homologies (structures ayant même fonction et même origine embryonnaire). On reconstruit donc une histoire, une généalogie. Remarque : le critère temporel est donc présent dans cette systématique. Les taxa dérivent les uns des autres. On regroupe toutes les espèces qui ont le même ancêtre. La paléontologie devient un outil intéressant dans ce cadre, mais pas suffisant à lui seul.
Cette systématique utilise aussi des caractères, mais ne les polarise pas, elle les hiérarchise en les regroupant en fonction de théories évolutives (par exemple : théorie évolutive de la forme, etc.). Cette systématique est surtout utilisée aujourd'hui par les botanistes.
|
Phylogramme simplifié des Monocotyledones
d'après W.J. Hahn ( http://tolweb.org/tree?group=Monocotyledons&contgroup=Angiosperms) |
V. - Petit glossaire
(attention, chaque classification a sa propre terminologie, mais utilise parfois les mêmes termes pour des notions différentes, ce glossaire est loin d'être complet mais est largement suffisant pour suivre le propos).
APOMORPHE : Etat d'un caractère quand celui-ci est dérivé, c’est-à-dire évolué par rapport à l’état plésiomorphe (ou primitif). C’est une notion relative, pas absolue ; un état de caractère ne peut être apomorphe (ou plésiomorphe) que par rapport à un autre état.
APOMORPHIE : Caractère apomorphe présent dans un taxon donné.
AUTAPOMORPHE : Lors d'une comparaison entre groupes-frères, c'est l'état dérivé d'un caractère propre à l'un des deux taxons.
AUTAPOMORPHIE : Caractère nouveau apparu dans un seul des groupes frères.
CARACTÈRE : Attribut d'un organisme utilisé pour le reconnaître, le différencier ou le classer.
CLADE sensu Simpson (1961) (κλαδοσ, branche). Groupe d'organismes vivants qui ont une origine génétique commune et qui satisfont a minima le critère de monophylie sensu Simpson (1961).
CLADE sensu Wiley & al. (1991) (κλαδοσ, branche). Unité monophylétique sensu Hennig (1966) délimitable de quelque niveau que ce soit.
CLADISME Synonyme de systématique phylogénétique. Ecole à la suite de Hennig, fondant la classification sur la proche parenté entre clades.
CLADOGENESE : Processus de division d'une espèce mère en deux espèces filles.
CLADOGRAMME Schéma exprimant les relations de proche parenté entre taxa construit à partir de l'analyse cladistique : les points intersections (noeuds) sont définis par des synapomorphies. Contrairement au phylogramme, les points de branchement ne reflètent pas les taux de divergence.
CONVERGENCE Ressemblance apparue indépendamment dans différents taxa. La convergence partagée par différents taxa n'est pas héritée de l'espèce ancestrale de ces taxa.
ESPECE POLYTYPIQUE : Espèce composée de populations présentant des phénotypes distincts bien que les individus des différentes populations conservent la capacité de s'accoupler en eux.
GRADE : Taxon souvent paraphylétique et parfois polyphylétique qui est supposé représenter un certain niveau de progrès évolutif, d'organisation ou d'adaptation.
HOLOPHYLETIQUE : Dans la terminologie de la classification évolutionniste un groupe est qualifié comme holophylétique s'il comprend seulement une espèce-mère (stem species) et toutes les espèces (taxa) descendant de celle-ci. Syn. : monophylétique sensu Hennig (1966).
HOLOPHYLIE : Dans la terminologie de la classification évolutionniste, il s'agit d'un cas particulier de monophylie sensu Simpson (1961) où la division est verticale de telle sorte que le groupe comprend seulement une espèce-mère (stem species) et toutes les espèces (taxa) qui descendent de celle-ci. Syn. : monophylie sensu Hennig (1966).
HOMOLOGIE : Partage d'un même caractère par différentes espèces en raison d'une ascendance commune.
HOMOPLASIE : Partage d'un même caractère par différentes espèces dû à la convergence et la réversion et non pas en raison d'une ascendance commune.
MONOPHYLETIQUE sensu Simpson (1961) : Dans la terminologie de la classification évolutionniste, un groupe est dit monophylétique sensu Simpson (1961) s'il provient d'une ou plusieurs lignées appartenant à un seul taxon de même rang ou d'un rang inférieur.
MONOPHYLETIQUE sensu Hennig (1966) : Dans la terminologie de la systématique phylogénétique, un groupe est dit monophylétique sensu Hennig (1966) s'il comprend seulement une espèce-mère (stem species) et toutes les espèces (taxa) qui descendent de celle-ci. Syn. : holophylétique.
MONOPHYLIE sensu Simpson (1961) : Dans la terminologie de la classification évolutionniste : L'état d'un groupe qui provient d'une ou plusieurs lignées appartenant à un seul taxon de même rang ou de rang inférieur.
MONOPHYLIE sensu Hennig (1966) : Dans la terminologie de la systématique phylogénétique : L'état d'un groupe qui comprend seulement une espèce-mère (stem species) et toutes les espèces (taxa) qui descendent de celle-ci. Syn. : holophylie.
MONOTHETIQUE : Un groupe est monothétique lorsqu'il peut être défini par la possession d'un seul caractère ou d'un ensemble restreint de caractères nécessaires et suffisants pour faire partie de ce groupe. Ant. : polythétique.
MONOTYPIQUE : Un genre (taxon) est dit monotypique lorsqu'il a été établi qu'à partir de l'existence attestée d'une seule espèce (taxon). Une espèce est dite monotypique si elle ne comprend pas de sous-espèces.
PARALLELOPHYLIE : Cas hypothétique de monophylie sensu Simpson (1961) où les synapomorphies et/ou les autapomorphies ne sont pas apparues en raison d'un ancêtre commun récent immédiat mais plutôt en raison de la transmission de gènes appartenant à un ancêtre commun lointain.
PARALLELISME : Il s'agit d'un cas particulier de convergence où la ressemblance est apparue indépendamment dans différents taxa proches parents.
PARAPHYLETIQUE sensu Simpson : Un groupe monophylétique est dit paraphylétique sensu Simpson lorsqu'il ne comprend pas toutes les espèces (taxa) qui sont issues d'une ou plusieurs espèces du groupe, le regroupement des lignées évolutives résultant d'une division horizontale.
PARAPHYLETIQUE sensu Wiley & al. (1991) : Un groupe artificiel est dit paraphylétique sensu Wiley & al. (1991) lorsqu'il ne comprend pas toutes les espèces (taxa) qui sont issues d'une ou plusieurs espèces du groupe, la division ayant été effectuée horizontalement pour tenir compte des symplésiomorphies.
PARAPHYLIE sensu Simpson (1961) : Dans la terminologie de la classification évolutionniste : Il s'agit d'un cas particulier de monophylie sensu Simpson (1961) où le regroupement des lignées évolutives résulte d'une division horizontale. Par conséquent, ce groupe ne comprend pas toutes les espèces (taxa) qui sont issues d'une ou plusieurs espèces du groupe.
PARAPHYLIE sensu Wiley & al. (1991) : Dans la terminologie de la systématique phylogénétique : L'état d'un groupe qui ne comprend pas toutes les espèces (taxa) qui sont issues d'une ou plusieurs espèces du groupe, la division ayant été effectuée horizontalement pour tenir compte des symplésiomorphies. Ce type de taxon est jugé artificiel car il n'est pas monophylétique sensu Hennig (1966).
PHENOTYPE : Ensemble des caractères d'un individu. Il correspond à la réalisation du génotype (globalité des gènes) sous l'influence de l'environnement et suite aux événements vécus par l'individu.
PLESIOMORPHE : Dans une série de transformations d'un caractère, se dit de l'état initial du caractère (synonyme de primitif). C'est une notion relatif (cf. Ant. Apomorphe)
POLYPHILIE : Dans la terminologie de la systématique phylogénétique : L'état d'un groupe qui comprend des espèces (taxa) qui proviennent de deux ou plusieurs espèces ancestrales différentes. Le regroupement est le fait du partage d'une ou plusieurs convergences. Ce type de taxon est jugé artificiel car il n'est pas monophylétique sensu Hennig (1966).
POLYTHETIQUE : Un groupe est dit polythétique lorsqu'il ne peut pas être défini par un caractère ou un ensemble restreint de caractères nécessaires et suffisants pour faire partie de ce groupe. Ainsi, une espèce appartient à un groupe polythétique donné plutôt à qu'à un autre parce qu'elle partage avec les membres de ce groupe un plus grand nombre de caractères qu'avec ceux des autres groupes. Ant. : monothétique.
SYMPLESIOMORPHIE : Le partage de caractére(s) plésiomorphe(s) ou ancien(s).
SYNAPOMORPHIE : Le partage de caractére(s) apomorphe(s) ou dérivé(s).
SYSTEMATIQUE : L'étude scientifique de la diversité des organismes vivants (s.l.)et des relations qu'ils entretiennent entre eux.
TAXON : (des taxons ou des taxa). Groupe d'organismes reconnu en tant qu'unité formelle à chacun des niveaux de la classification
VI. - Bibliographie succinte
DARLU P. & TASSY P. (1993) La reconstruction phylogénétique : concepts et méthodes. Masson, Paris.
HENNIG W. (1950) Grundzüge einer Theorie der Phylogenetischen Systematik. Berlin, Deutcher Zentralverlag.
ROSS H.H. (1973) Biological systematics. Addison-Wesley, Reading, Mass.
SIMPSON G.G. (1961) Principles of animal taxonomy. N.Y., Columbia Univ. Press.
SMITH A.B. (1988) Phylogenic relationship, divergence times and rates of molecular evolution for Camarodont sea urchins. Mol. Biol. Evol. 5, (4), p.345-365.
SNEATH P.H. & SOKAL R.R. (1973) Numerical taxonomy. The principles and practice of numerical classification. San Francisco, Freeman and Co.
SOBER E. (1991) Reconstructing the past : parsimony, evolution and inference. Cambridge, Mass. Inst. of Tech. press.
WILEY E.O. (1981) Phylogenetics. The theory and practice of phylogenetic systematics. Wiley, N.Y.
WILEY E.O., SIEGEL-CAUSEY D., BROOKS D.R. & FUNK V.A. (1991) The compeat Cladist : A primer of phylogenetic procedures. Univ. Kansas Mus. of Nat. Hist. (19)